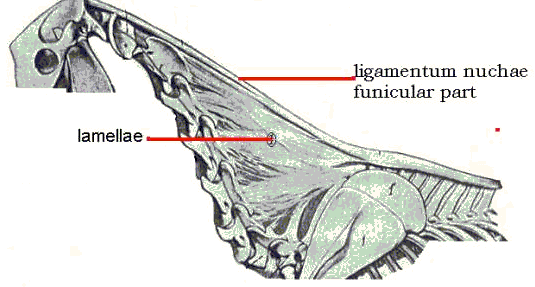
Neck Movement (Kinematics) of the Horseby Observations were made on the movement of the cervical vertebrae of the horse. There appears to be little relevant literature other than that of Demeter (1916), Smith (1921), Rooney (1969), and Clayton and Townsend (1989). The anatomy is described in Ellenberger-Baum (1943) and other veterinary anatomical texts. This investigation was anatomical and mechanical and restricted to the movement of the head and neck in the sagittal plane. Additional studies are in progress on lateral and rotational movements. The head/neck system was dissected and manipulated in six fresh carcasses, removing the extrinsic muscles attached to the cervical vertebrae but leaving the head, intrinsic muscles, and ligamentum nuchae intact, Figure 1. (After Ellenberger and Baum)
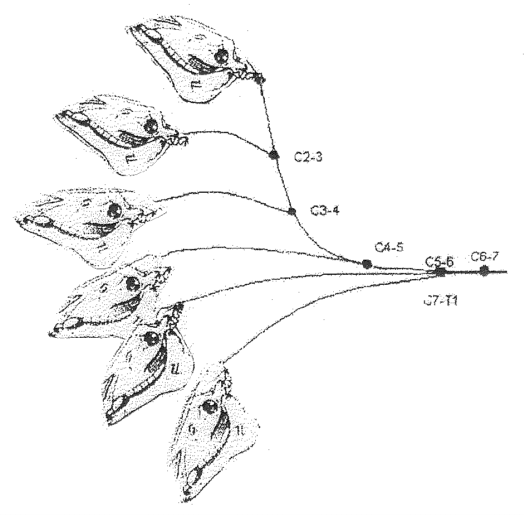
Results In the ventroflexed – grazing - position the neck is concave ventrally. When fully raised - dorsiflexed - the neck is concave dorsally. In the usual standing position, between ventroflexed and dorsiflexed, the cervical vertebral column assumes a sigmoid, S-shape. As the neck moves from dorsiflexion through the S-shaped position into ventroflexion, the cervical vertebrae move smoothly in series: C2-3, C3-4, C4-5, C5-6, C6-7, C7-T1, Figure 2.
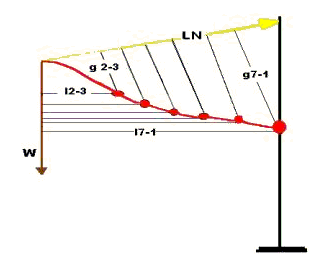
Movement at the atlantoccipital joint does not effect neck movements and is not considered here. There is little or no movement in the sagittal plane between the atlas and axis (C1-2) and very little movement between C2-3 (Rooney 1969). The mechanics may be described as follows, Figure 3.
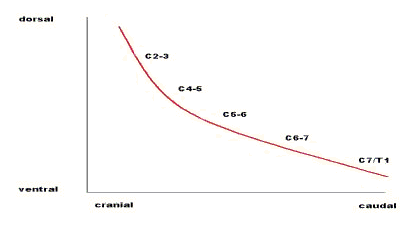
The neck is considered as a flexible, jointed, cantilever beam (as, also, in the model below) with the head as the weight (W) on the free end of the beam. This is not strictly true but is sufficiently accurate for present purposes. W acts around distances, l, to produce counterclockwise rotations (moments)at each successive intervertebral center from C2-3 to C7-T1. In Figure 3, l2-3 is the moment arm, l, perpendicular to the line of action of W to the center of rotation at cervical intervertebral joint 2-3. Similarly, l7-1 is the moment arm of W to the center at C7-T1. The other moment arms are not labeled in order to avoid clutter in the figure. These moments are balanced by clockwise moments produced by the tensile force (LN) in the funicular ligamentum nuchae together with the tonically active dorsal muscles of the neck acting over the distances, g. The notation is as given above. The value of W is constant. At any given instant or static position, LN is constant, and we can write the moment equilibrium equation: Wl - LNg = 0 (Obviously, this is oversimplified; while W and LN are constant, the values of l and g are different at each intervertebral joint. The equation, then, can be considered as an average with the values of l and g being averaged.) In order for the head and neck to ventroflex, move downward, LN decreases, so that Wl is effectively greater than LNg. That is, either the dorsal cervical muscles relax and/or the ventral muscles contract (the ligamentum nuchae is, of course, a passive structure). In Figure3 it is apparent that the moment produced by LN is greatest at C7-T1 and decreases cranially; that is, the g at each cervical intervertebral junction is less as one moves cranially. As Wl causes downward movement, then, the resisting moments of LN are least cranially, increasing caudally to C7-T1. Therefore, cervical vertebrae ventroflex in series as described. The system operates in reverse as the head and neck are raised. From the ventroflexed position rotation begins at C7-T1 and continues in series to C2-3 as the neck passes through the sigmoid position into dorsiflexion. As LN increases-by muscle action- those joints with larger LN moments will move first just as those joints with smaller LN moments moved first when the neck moved in the opposite direction. There is, then, a locus of centers of rotation as the neck moves up and down, that locus having the form of an exponential curve, Figure 4.That is, as the neck moves there is not one center of rotation, such as the axle of a wheel, but a series of centers or rotation, one after the other.
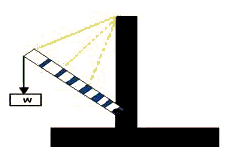
The model of Figure 5 is readily constructed and helpful in visualizing the action of the neck with and without the ligamentum nuchae. Blocks of wood representing the vertebrae are glued to pieces of rubber or plastic foam representing the intervertebral disks. A rubber band serves as the funicular ligamentum nuchae. Without the rubber band the model simply bends downward, rotating around the center formed by the last block and the support (C7-T1). With the rubber band in place increasing W bends the neck into the S-shaped sigmoid curve continuing into ventroflexion just as in the anatomical specimen. In life, of course, W is constant; increasing W in the model or the anatomical specimen is equivalent to decreasing dorsiflexing muscular action and increasing ventroflexing muscular action as already noted.
The lamellar ligamentum nuchae are tense when the neck is in the sigmoid position and remain tense as the neck is dorsiflexed, beginning to loosen only as the limit of dorsiflexion is approached. This limit in the anatomical specimen in vitro appears to be beyond what is possible and/or usual in the live horse. As the neck ventroflexes the lamellae tighten and collapse upon themselves. Demeter (1916) believed that the lamellae caused the S-shape, but that shape is caused by the funicular part of the ligamentum nuchae alone as demonstrated by removing the lamellae. The S-shape also appears in the model without lamellar elements. The action of the lamellae can be included in the model with additional rubber bands as indicated by the dotted lines in Figure 5. The lamellae, then, are tense and assisting the funicular ligamentum nuchae and dorsal musculature in supporting the head and neck in all positions except marked dorsiflexion. Discussion Sagittal movement of the neck is controlled by the ligamentum nuchae and those cervical muscles acting in parallel with the ligamentum nuchae. Because of the passive, energy-sparing ligamentum nuchae, less muscular work is required to maintain the head and neck in any given position. A basic, tonic muscular contribution is, however, necessary as indicated by the lack of muscular tonus in seriously ill or debilitated animals which stand with the head and neck more ventroflexed than in the usual standing position. Clayton and Townsend (1989) found that the dorsoventral movement of the cervical vertebrae in vitro was greatest at C7-T1 and decreased to C1-2. This is certainly true for the neck deprived of the action of muscles and the ligamentum nuchae (Rooney 1969). Resistance to movement is provided by a combination of muscle, ligaments, and intervertebral disks. With muscle and ligamentum nuchae removed, as in the Clayton and Townsend study, there are only intervertebral disks and the fibrous joint capsules, and the neck moves like a rod, a simple cantilever beam, rotating around the C7-T1 center as it moves from the dorsiflexed to the ventroflexed position. When the ligamentum nuchae is intact, the series rotation described herein pertains. Smith (1921) mentioned the function of the funicular ligamentum nuchae. He believed it assisted the muscles in keeping the head extended as, for example, when grazing. He also said that shortening of the ligament was responsible for the dorsiflexion (opisthotonus) of the head/neck after death. This is not the case since severing the ligament does not release such dorsiflexion; rigor mortis of the dorsal cervical muscles causes opisthotonus after death. Smith also believed that the ligamentum nuchae did not have an important role in supporting the head since the head did not droop down when the funicular part was divided. Since the position of the head and neck is a function of both muscle and ligament action, the muscles together with the lamellar part of the ligamentum nuchae support the head and neck when the funicular part of the ligament is severed. It is clear from dissected specimens with the muscle removed and the head and neck positioned as in life that the ligamentum nuchae plays an important passive role. Bibliography Clayton, H M and Townsend, H G G (1989) Kinematics of the cervical spine of the adult horse. Equine Veterinary Journal 21: 189-192. Return to Dr. Rooney's home page.
|

Trackback URL : 이 글에는 트랙백을 보낼 수 없습니다
Trackback RSS : http://www.fallight.com/rss/trackback/1377
Trackback ATOM : http://www.fallight.com/atom/trackback/1377







